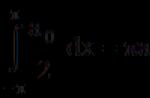Рибонуклеиновая кислота представляет собой сополимер пуриновых и пиримидиновых рибонуклеотидов, соединенных друг с другом, как и в ДНК, -фосфодиэфирными мостиками (рис. 37.6). Хотя эти два вида нуклеиновых кислот имеют много общего, по ряду признаков они отличаются друг от друга.
1. У РНК углеводным остатком, к которому присоединены пуриновые или пиримидиновые основания и фосфатные группы, является рибоза, а не 2-дезоксирибоза (как у ДНК).
2. Пиримидиновые компоненты РНК отличаются от таковых у ДНК. В состав РНК, как и в состав ДНК, входят нуклеотиды аденина, гуанина и цитозина. В то же время РНК (за исключением некоторых специальных случаев, на которых мы остановимся ниже) не содержит тимина, его место в молекуле РНК занимает урацил.
3. РНК - одноцепочечная молекула (в отличие от ДНК, имеющей двухцепочечную структуру), однако при наличии в цепи РНК участков с комплементарной последовательностью (противоположной полярности) единичная цепь РНК способна сворачиваться с образованием так называемых «шпилек», структур, имеющих двухспиральные характеристики (рис. 37.7).

Рис. 37.6. Фрагмент молекулы рибонуклеиновой кислоты (РНК), в котором пуриновые и пиримидиновые основания- аденин (А), урацил (U), цитозин (С) и гуанин (-удерживаются фосфодиэфирным остовом, соединяющим рибозильные остатки, связанные N-гликозидной связью с соответствующими нуклеиновыми основаниями. Обратите внимание: цепь РНК обладает определенной направленностью, на которую указывают 5- и З-концевые фосфатные остатки.
4. Так как молекула РНК представляет собой одиночную цепь, комплементарную только одной из цепей ДНК, содержание в ней гуанина не обязательно равно содержанию цитозина, а содержание аденина не обязательно равно содержанию урацила.
5. РНК может быть гидролизована щелочью до 2, З-циклических диэфиров мононуклеотидов; в роли промежуточного продукта гидролиза выступает 2, У, 5-триэфир, который не образуется при щелочном гидролизе ДНК из-за отсутствия у последней 2-гидроксильных групп; щелочная лабильность РНК (сравнительно с ДНК) является полезным свойством как для диагностических, так и для аналитических целей.
Информация, содержащаяся в одноцепочечной РНК, реализуется в виде определенной последовательности пуриновых и пиримидиновых оснований (т. е. в первичной структуре) полимерной цепи. Эта последовательность комплементарна кодирующей цепи гена, с которой «считывается» РНК. Вследствие комплементарности молекула РНК способна специфически связываться (гибридизоваться) с кодирующей цепью, но не гибридизуется с некодирующей цепью ДНК. Последовательность РНК (за исключением замены Т на U) идентична последовательности некодирующей цепи гена (рис. 37.8).
Биологические функции РНК
Известно несколько видов РНК. Почти все они непосредственно вовлечены в процесс биосинтеза белка. Молекулы цитоплазматической РНК, выполняющие функции матриц белкового синтеза, называются матричными РНК (мРНК). Другой вид цитоплазматической РНК-рибосомная РНК (рРНК) - выполняет роль структурных компонентов рибосом (органелл, играющих важную роль в синтезе белка). Адапторные молекулы транспортных РНК (тРНК) участвуют в трансляции (переводе) информации мРНК в последовательность аминокислот в белках.
Значительная часть РНК-первичных транскриптов, образующихся в эукариотических клетках, включая и клетки млекопитающих, - подвергается деградации в ядре и не играет какой-либо структурной или информационной роли в цитоплазме. В культивируемых

Рис. 37.7. Вторичная структура молекулы РНК типа «петли со стеблем» («шпилька»), возникающая вследствие внутримолекулярного образования водородных связей между комплементарными парами нуклеиновых оснований.
клетках человека обнаружен класс малых ядерных РНК которые непосредственно не участвуют в синтезе белка, но могут оказывать влияние на процессинг РНК и общую «архитектуру» клетки. Размеры этих относительно небольших молекул варьируют, последние содержат от 90 до 300 нуклеотидов (табл. 37.3).
РНК является основным генетическим материалом у некоторых вирусов животных и растений. Некоторые РНК-содержащие вирусы никогда не проходят стадию обратной транскрипции РНК в ДНК. Однако для большинства известных вирусов животных, таких, как ретровирусы, характерна обратная транскрипция их РНК-генома, направляемая РНК-зависимой ДНК-полимеразой (обратной транскриптазой) с образованием двухспиральной ДНК-копии. Во многих случаях образующийся двухспиральный ДНК-транскрипт встраивается в геном и в дальнейшем обеспечивает экспрессию генов вируса, а также наработку новых копий вирусных РНК-геномов.
Структурная организация РНК
Во всех эукариотических и прокариотических организмах существуют три основных класса молекул РНК: информационная (матричная или мессенджер) РНК (мРНК), транспортная (тРНК) и рибосомная (рРНК). Представители этих классов отличаются друг от друга размерами, функциями и стабильностью.
Информационная (мРНК) - наиболее гетерогенный в отношении размеров и стабильности класс. Все представители этого класса служат переносчиками информации от гена к белок-синтезирующей системе клетки. Они выполняют роль матриц для синтезируемого полипептида, т. е. определяют аминокислотную последовательность белка (рис. 37.9).
Информационные РНК, особенно эукариотические, обладают некоторыми уникальными структурными особенностями. 5-Конец мРНК «кэпирован» 7-метилгуанозинтрифосфатом, присоединенным к 5-гидроксилу соседнего 2-0-метилрибонуклеозида через остаток трифосфата (рис. 37.10). Молекулы мРНК часто содержат внутренние остатки 6-метиладенина и 2-0-метилированные рибонуклеотиды. Хотя смысл «кэпирования» до конца еще не выяснен, можно предположить, что образующаяся структура 5-конца мРНК используется для специфического узнавания в системе трансляции. Синтез белка начинается на 5"-(кэпированном) конце мРНК. Другой конец большинства молекул мРНК (З-конец) содержит полиаденилатную цепочку из 20-250 нуклеотидов. Специфические функции этого окончательно не установлены. Можно предполагать, что данная структура отвечает за поддержание внутриклеточной стабильности мРНК. Некоторые мРНК, включая гистоновые не содержат poly (А). Наличие poly (А) в структуре мРНК используется для отделения от других видов РНК посредством фракционирования тотальной РНК на колонках с oligo (Т), иммобилизованным на твердом носителе типа целлюлозы. Связывание мРНК с колонкой происходит за счет комплементарных взаимодействий poly (А)-«хвоста» с иммобилизованным oligo (Т).
Рис. 37.8. Последовательность гена и его РНК-транскрипта. Показаны кодирующая и некодирующая цепи, и отмечена их полярность. РНК-транскрипт, имеющий полярность комплементарен кодирующей цепи (с полярностью 3 - 5) и идентичен по последовательности (за исключением замен Т на U) и полярности некодирующей цепи ДНК.

Рис. 37.9. Экспрессия генетической информации ДНК в форме мРНК-транскрипта и последующая трансляция при участии рибосом с образованием специфической молекулы белка.
(см. скан)
Рис. 37.10. Структура «кэпа», находящегося на 5-конце большинства эукариотических матричных РНК 7-метилгуанозинтрифосфат присоединяется к 5-концу мРНК. на котором обычно находится 2-О-метилпуриновый нуклеотид.
В клетках млекопитающих, включая клетки человека, зрелые молекулы мРНК, находящиеся в цитоплазме, не являются полной копией транскрибируемого участка гена. Образующийся в результате транскрипции полирибонуклеотид представляет собой предшественник цитоплазматической мРНК, перед выходом из ядра он подвергается специфическому процессингу. Непроцессированные продукты транскрипции, обнаруживаемые в ядрах клеток млекопитающих, образуют четвертый класс молекул РНК. Такие ядерные РНК очень гетерогенны и достигают значительных размеров. Молекулы гетерогенных ядерных РНК могут иметь молекулярную массу более , в то время как молекулярная масса мРНК обычно не превышает 2106. подвергаются процессингу в ядре, и образующиеся зрелые мРНК поступают в цитоплазму, где служат матрицей для биосинтеза белка.
Молекулы транспортных РНК (тРНК) обычно содержат около 75 нуклеотидов. Молекулярная масса таких молекул составляет . тРНК также формируются в результате специфического процессинга соответствующих молекул-предшественников (см. гл. 39). Транспортные тРНК выполняют функцию посредников в ходе трансляции мРНК. В любой клетке присутствуют не менее 20 видов молекул тРНК. Каждый вид (иногда несколько видов) тРНК соответствует одной из 20 аминокислот, необходимых для синтеза белка. Хотя каждая специфическая тРНК отличается от других нуклеотидной последовательностью, все они имеют и общие черты. Благодаря нескольим внутрицепочечным комплементарным участкам, все тРНК обладают вторичной структурой, получившей название «клеверный лист» (рис. 37.11).
Молекулы всех видов тРНК имеют четыре основных плеча. Акцепторное плечо состоит из «стебля» спаренных нуклеотидов и заканчивается последовательностью ССА Именно через У-гидроксильную группу аденозильного остатка происходит связывание с карбоксильной группой аминокислоты. Остальные плечи тоже состоят из «стеблей», образованных комплементарными парами оснований, и петель из неспаренных оснований (рис. 37.7). Антикодоновое плечо узнает нуклеотидный триплет или кодон (см. гл. 40) в мРНК. D-плечо названо так из-за наличия в нем дигидроуридина, -плечо названо по последовательности Т-псевдоуридин-С. Дополнительное плечо представляет собой наиболее вариабельную структуру и служит основой классификации тРНК. тРНК класса 1 (75% от общего их числа) обладают дополнительным плечом длиной 3-5 пар оснований. Дополнительное плечо у тРНК-молекул класса 2 состоит из 13-21 пар оснований и часто включает неспаренную петлю.

Рис. 37.11. Структура молекулы аминоацил-тРНК, к 3-ССА-концу которой присоединена аминокислота . Указаны внутримолекулярные водородные связи и расположение антикодонового, ТТС- и дигидроурацилового плеч. (From J. D. Watson. Molecular biology of the Gene 3rd, ed.. Copyright 1976, 1970, 1965 by W. A. Benjamin, Inc., Menlo Park Calif.)
Вторичная структура, определяемая системой комплементарных взаимодействий нуклеотидных оснований соответствующих плеч, характерна для всех видов Акцепторное плечо содержит семь пар оснований, -плечо - пять пар оснований, плечо D - три (или четыре) пары оснований.
Молекулы тРНК весьма стабильны у прокариот и несколько менее стабильны у эукариот. Обратная ситуация характерна для мРНК, которая довольно нестабильна у прокариот, а у эукариотических организмов обладает значительной стабильностью.
Рибосомная РНК. Рибосома - это цитоплазматическая нуклеопротеиновая структура, предназначенная для синтеза белка по мРНК-матрице. Рибосома обеспечивает специфический контакт в результате которого и происходит трансляция нуклеотидной последовательности, считанной с определенного гена, в аминокислотную последовательность соответствующего белка.
В табл. 37.2 представлены компоненты рибосом млекопитающих, имеющих молекулярную массу 4,210 6 и скорость седиментации (единиц Сведберга). Рибосомы млекопитающих состоят из двух нуклеопротеиновых субъединиц - большой с
Таблица 37.2. Компоненты рибосом млекопитающих

молекулярной массой (60S), и малой, имеющей молекулярную массу (40S). 608-субъединица содержит 58-рибосомную РНК (рРНК), 5,8S-pPHK и 28S-pPHK, а также более 50 различных полипептидов. Малая, 408-субъединица включает единственную 18S-pPHK и около 30 полипептидных цепей. Все рибосомные РНК, за исключением 5S-PHK, имеют общего предшественника-45S-PHK, локализованную в ядрышке (см. гл. 40). У молекулы 5S-PHK предшественник собственный. В ядрышке происходит упаковка высокометилированных рибосомных РНК с рибосомными белками. В цитоплазме рибосомы достаточно устойчивы и способны осуществлять большое число циклов трансляции.
Небольшие стабильные РНК. В эукариотических клетках обнаружено большое число дискретных, высококонсервативных, небольших и стабильных молекул РНК. Большинство РНК этого типа обнаруживаются в составе рибонуклеопротеинов и локализованы в ядре, цитоплазме или одновременно в обоих компартментах. Размеры этих молекул варьируют от 90 до 300 нуклеотидов, содержание их - 100000-1000000 копий на клетку.
Малые ядерные нуклеопротеиновые частицы (часто называемые snurps - от англ. small nuclear ribonucleic particles), вероятно, играют существенную роль в регуляции экспрессии генов. Нуклеопротеиновые частицы типа U7, по-видимому, участвуют в формировании З-концов гистоновых мРНК. Частицы , вероятно, необходимы для полиаденилирования, a - для удаления интронов и процессинга мРНК (см. гл. 39). Табл. 37.3. суммирует некоторые характеристики небольших стабильных РНК.
Таблица 37.3. Некоторые виды небольших стабильных РНК, обнаруженные в клетках млекопитающих

ЛИТЕРАТУРА
Darnell J. et al. Molecular Cell Biology, Scientific American Books, 1986.
Hunt T. DNA Makes RNA Makes Protein, Elsevier, 1983. Lewin B. Genes, 2nd ed., Wiley, 1985.
Rich A. et al. The chemistry and biology of left-handed Z-DNA, Annu. Rev. Biochem., 1984, 53, 847.
Turner P. Controlling roles for snurps, Nature, 1985, 316, 105. Watson J. D. The Double Helix, Atheneum, 1968.
Watson J. D., Crick F.H.C. Molecular structure of nucleic acids. Nature, 1953, 171, 737.
Zieve G. W. Two groups of small stable RNAs, Cell, 1981, 25, 296.

Рис. 17. Составные части зрелой мРНК
1. «Колпачок» (кэп) – нуклеотидная последовательность на 5′-конце мРНК, состоящая из одного-четырех модифицированных нуклеотидов. Такая структура защищает 5′-конец мРНК от действия экзонуклеаз.
2. 5′-нетранслируемый участок – последовательность из нескольких десятков нуклеотидов, которые комплементарны нуклеотидам рРНК, входящим в состав малой субъединицы рибосомы. За счет этого 5′-нетранслируемый участок выполняет функцию первичного связывания мРНК с рибосомой, но сам не транслируется.
3. Инициирующий кодон – кодон, с которого начинается трансляция мРНК. Во всех мРНК он один и тот же – АУГ (кодирует метионин).
4. Кодирующая часть содержит информацию о последовательности аминокислот в белке. В зрелой мРНК она лишена интронов – вставочных некодирующих последовательностей, т. е. имеется непрерывная последовательность смысловых кодонов, которая должна считываться в направлении 5′®3′.
5. Терминирующий кодон – один из трех «бессмысленных» кодонов: УАА, УАГ или УГА.
6. 3′-нетранслируемый участок содержит последовательности нуклеотидов, которые называют элементами нестабильности мРНК. Определенные белки клетки узнают эти последовательности, связываются с ними и стабилизируют мРНК.
7. Поли (А)-фрагмент содержит от 50 до 400 адениловых нуклеотидов. Эти фрагменты отсутствуют в молекулах гистоновых мРНК. Полагают, что поли (А)-фрагмент имеет отношение к регуляции продолжительности жизни мРНК. Согласно одной из гипотез, после того как очередная рибосома заканчивает трансляцию мРНК, от поли (А)-фрагмента отщепляются 10-15 нуклеотидов. Когда данный фрагмент исчерпывается, начинает разрушаться кодирующая часть мРНК (если отсутствует 3′-нетранслируемый участок).
Общее количество нуклеотидов в мРНК составляет обычно несколько тысяч. При этом на кодирующую часть приходится лишь 60-70% нуклеотидов.
 Рис. 18. Структура тРНК (на примере тРНК Phe)
Рис. 18. Структура тРНК (на примере тРНК Phe)
|
 Транспортные РНК (тРНК)
– короткие молекулы (70-90 нуклеотидов), главной функцией которых является присоединение аминокислот и перенос их в белоксинтезирующий аппарат клетки. Количество различных тРНК в клетке – несколько десятков: от одного до шести видов для каждой из 20 аминокислот. Виды тРНК, способные связывать одну и ту же аминокислоту, называются изоакцепторными. Специфичность тРНК обознается верхним индексом, например тРНК Ала.
Транспортные РНК (тРНК)
– короткие молекулы (70-90 нуклеотидов), главной функцией которых является присоединение аминокислот и перенос их в белоксинтезирующий аппарат клетки. Количество различных тРНК в клетке – несколько десятков: от одного до шести видов для каждой из 20 аминокислот. Виды тРНК, способные связывать одну и ту же аминокислоту, называются изоакцепторными. Специфичность тРНК обознается верхним индексом, например тРНК Ала.
Среди нуклеотидов тРНК высоко содержание минорных нуклеотидов (около 10%). Благодаря образованию участков вторичной структуры, цепь тРНК имеет характерную форму «клеверного листа» (рис. 18).
В этой структуре четыре двухцепочечных и пять одноцепочечных участков. Поскольку минорные нуклеотиды, как правило, не способны к комплементарным взаимодействиям, они содержатся в основном в одноцепочечных локусах.
Для одноцепочечных участков приняты следующие названия:
· акцепторная ветвь – участок на 3′-конце тРНК из четырех нуклеотидов, место присоединения аминокислоты;
· антикодоновая петля – участок из 7 нуклеотидов в середине цепи; три из них выполняют функцию антикодона, который комплементарно взаимодействует с соответствующим кодоном мРНК;
· дигидроуридиловая, псевдоуридиловая и не всегда имеющаяся добавочная петли способствуют формированию специфичной для данной тРНК третичной структуры.
Особенность тРНК – наличие стабильной третичной структуры. Четыре двухцепочечных участка, попарно сближаясь, образуют примерно два витка двойной спирали, расположенные почти перпендикулярно друг другу – так, что молекула приобретает Г-образную форму (рис. 16Б).
 Рибосомные РНК
являются структурной основой для формирования субъединиц рибосом. Среди азотистых оснований в рРНК выше, чем обычно, содержание гуанина и цитозина. Встречаются также минорные азотистые основания, но не столь часто, как в тРНК – примерно 1%. Во вторичной структуре рРНК много двухцепочечных участков и петель (рис. 19).
Рибосомные РНК
являются структурной основой для формирования субъединиц рибосом. Среди азотистых оснований в рРНК выше, чем обычно, содержание гуанина и цитозина. Встречаются также минорные азотистые основания, но не столь часто, как в тРНК – примерно 1%. Во вторичной структуре рРНК много двухцепочечных участков и петель (рис. 19).
Рис. 19. Структура 16S-рРНК прокариотической и 18S-рРНК эукариотической клеток
 Рибосомные РНК, а также образуемые ими субъединицы рибосом принято обозначать по их константе седиментации (S). Седиментация (от лат. sedimentum
– осадок) – оседание частиц, взвешенных в жидкости, при центрифугировании. Под действием центробежных сил, намного превосходящих силу тяжести, даже сравнительно небольшие макромолекулы, такие как тРНК, разделяются и распределяются в строгом соответствии со своими размерами. Измерение коэффициента седиментации макромолекулярных комплексов обычно используют для определения их общей массы и количества входящих в их состав субъединиц.
Рибосомные РНК, а также образуемые ими субъединицы рибосом принято обозначать по их константе седиментации (S). Седиментация (от лат. sedimentum
– осадок) – оседание частиц, взвешенных в жидкости, при центрифугировании. Под действием центробежных сил, намного превосходящих силу тяжести, даже сравнительно небольшие макромолекулы, такие как тРНК, разделяются и распределяются в строгом соответствии со своими размерами. Измерение коэффициента седиментации макромолекулярных комплексов обычно используют для определения их общей массы и количества входящих в их состав субъединиц.
Рис. 20. Состав цитоплазматической рибосомы эукариот (в скобках указано
количество нуклеотидов в цепи рРНК)
Размеры молекул рРНК в рибосомных субъединицах у разных организмов варьируют (рис. 20), но сложная ее структура остается относительно постоянной. Кроме рРНК в состав рибосомы входит значительное число белков, но многие из них, по-видимому, не являются необходимыми для функционирования рибосом. Можно предположить, что именно молекулы РНК, а не белковые молекулы катализируют многие реакции, протекающие на рибосомах. Рибосомные белки при этом лишь усиливают функции рРНК.
Строение и функции РНК
| Наименование параметра | Значение |
| Тема статьи: | Строение и функции РНК |
| Рубрика (тематическая категория) | Дом |
Строение РНК – полимер, мономерами которого служат нуклеотиды. Три азотистых основания те же, что в составе ДНК (аденин, гуанин, цитозин ); четвертое - урацил - присутствует в молекуле РНК вместо тимина. Нуклеотиды РНК содержат вместо дизоксирибозы рибозу . В цепочке РНК нуклеотиды соединяются ковалентными связями между рибозой одного нуклеотида и остатком фосфорной кислоты другого.
В организме РНК находятся в виде комплексов с белками - рибонуклеопротеидов.
Известны 2 типа молекул РНК: 1) Двуцепочные РНК характерны для некоторых вирусов – служат для хранения и воспроизведения наследственной информации (выполняют функции хромосом). 2) У большинства клеток - одноцепочные РНК – осуществляют перенос информации об аминокислотной последовательности в белках от хромосомы к рибосоме.
Одноцепочечные РНК имеют пространственную организацию : за счёт взаимодействия азотистых оснований друг с другом, а также с фосфатами и гидроксилами сахарофосфатного остова происходит сворачивание цепи в компактную структуру типа глобулы. Функция: перенос от хромосомы к рибосомам информацию о последовательности АК в белках, которые должны синтезироваться.
Существует несколько типов одноцепочных РНК по выполняемой функции или месту нахождения в клетке:
1. Рибосомная РНК (рРНК) составляет основную часть РНК цитоплазмы (80-90 %). Размеры 3000-5000 пар нуклеотидов. Вторичная структура в виде двухспиральных шпилек. р-РНК является структурным компонентом рибосом - органоиды клетки, где происходит синтез белков. Рибосомы локализованы в цитоплазме, ядрышке, митохондриях, хлоропластах. Состоят из двух субъединиц – большой и малой. Малая субчастица состоит из одной молекулы рРНК и 33 молекул белков, большая субъединица - 3 молекулы рРНК и 50 белков. Белки рибосом выполняют ферментативную и структурную функции. Функции р-РНК: 1) структурный компонент рибосом – их целостность необходима для биосинтеза белков, 2) обеспечивают правильность связывания рибосомы с м-РНК, 3) 2) обеспечивают правильность связывания рибосомы с т-РНК.
2. Матричная (мРНК ) – 2-6 % от общего количества РНК. Состоит из участков: 1) цистроны – определяют последовательность АК в кодируемых ими белках, имеют уникальную последовательность нуклеотидов, 2) нетранслируемые области располагаются на концах молекулы, имеют общие закономерности нуклеотидного состава.
Кэп – особая структура на 5′ конце м-РНК - ϶ᴛᴏ 7-метилгуанозинтрифосфат, образуется ферментативным путем в процессе транскрипции. Функции кэпа : 1) предохраняет 5′ конец от расщепления экзонуклеазами, 2) используется для специфического узнавания м-РНК в процессе трансляции.
Прецистронный нетранслируемый участок – 3-15 нуклеотидов. Функция : обеспечение правильного взаимодействия 5′ конца м-РНК с рибосомой.
Цистрон: содержит инициирующий и терминирующий кодоны – осо-
бые последовательности нуклеотидов, отвечающие за начало и окончание передачи информации с данного цистрона.
Постцистронный нетранслируемый участок – находится на 3′ конце, содержит гексануклеотид (часто ААУААА) и цепочку из 20-250 адениловых нуклеотидов. Функция – поддержание внутриклеточной стабильности м-РНК.
3. Транспортные РНК (тРНК ) – 15 % от общей РНК, состоят из 70-93 пар нуклеотидов. Функция: перенос аминокислоты к месту синтеза белка, ʼʼузнаютʼʼ (по принципу комплиментарности) участок мРНК, соответствующий переносимой аминокислоте. Для каждой из 20 АК имеются специфические т-РНК (обычно более одной). Все т-РНК имеют сложную структуру, изображаемую в виде клеверного листа. Клеверный лист содержит 5 участков:
1) 3′ конец – акцепторная ветвь (сюда присоединяется эфирной связью остаток АК),
2) антикидоновая ветвь – располагается напротив акцепторного участка, состоит из трёх неспаренных (имеющих свободные связи) нуклеотидов (антикодон) и специфически спаривается (антипараллельно, комлиментарно) с кодоном м-РНК.
Кодон – набор из 3 нуклеотидов (триплет) в м-РНК, определяющий место данной аминокислоты в синтезируемой полипептидной цепи. Это единица генетического кода, с помощью которого в молекулах ДНК и РНК ʼʼзаписанаʼʼ вся генетическая информация.
3) Т-ветвь (псевдоурединовая петля - содержит псевдоуредин) – участок, присоединяющийся к рибосоме. 4) Д-ветвь (дегидроуреди6новая петля - содержит дегидроуредин) – участок, обеспечивающий взаимодействие с соответствующим аминокислоте ферментом аминоацил-тРНК-синтетазой. 5) Дополнительная малая ветвь. Функции пока не изучены.
4) Ядерные РНК (яРНК) – компонент ядра клеток. Низкополимерная, стабильная, роль которой пока неясна.
Все виды РНК синтезируются в клеточном ядре на матрице ДНК под действием ферментов полимераз . При этом образуется последовательность рибонуклеотидов, комплементарная последовательности дезоксирибо-нуклеотидов в ДНК - ϶ᴛᴏ процесс транскрипции.
Строение и функции РНК - понятие и виды. Классификация и особенности категории "Строение и функции РНК" 2017, 2018.
Время, в которое мы живем, отмечено потрясающими переменами, огромным прогрессом, когда люди получают ответы на все новые и новые вопросы. Жизнь стремительно движется вперед, и то, что еще совсем недавно казалось невозможным, начинает претворяться в жизнь. Вполне возможно, что представляется сегодня сюжетом из жанра фантастики, скоро тоже приобретет черты реальности.
Одним из важнейших открытий во второй половине двадцатого столетия стали нуклеиновые кислоты РНК и ДНК, благодаря которым человек приблизился к разгадкам тайн природы.
Нуклеиновые кислоты
Нуклеиновые кислоты - это органические соединения, обладающие высокомолекулярными свойствами. В их состав входят водород, углерод, азот и фосфор.
Они были открыты в 1869 году Ф. Мишером, который исследовал гной. Однако тогда его открытию не придали особого значения. Лишь позже, когда эти кислоты обнаружили во всех животных и растительных клетках, пришло понимание огромной их роли.
Существуют два вида нуклеиновых кислот: РНК и ДНК (рибонуклеиновые и дезоксирибонуклеиновые кислоты). Настоящая статья посвящена рибонуклеиновой кислоте, но для общего понимания рассмотрим также, что собой представляет ДНК.
Что такое
ДНК — это состоящая из двух нитей, которые соединены по закону комплементарности водородными связями азотистых оснований. Длинные цепи закручены в спираль, один виток содержит почти десять нуклеотидов. Диаметр двойной спирали составляет два миллиметра, расстояние между нуклеотидами - около половины нанометра. Длина одной молекулы порой достигает нескольких сантиметров. Длина ДНК ядра человеческой клетки составляет почти два метра.
В структуре ДНК содержится вся ДНК обладает репликацией, что означает процесс, в ходе которого из одной молекулы образуются две совершенно одинаковые - дочерние.
Как уже было отмечено, цепь складывается из нуклеотидов, состоящих, в свою очередь, из азотистых оснований (аденина, гуанина, тимина и цитозина) и остатка кислоты фосфора. Все нуклеотиды различаются азотистыми основаниями. Водородная связь возникает не между всеми основаниями, аденин, к примеру, может соединяться только с тимином или гуанином. Таким образом, адениловых нуклеотидов в организме столько же, сколько тимидиловых, а число гуаниловых равно цитидиловым (правило Чаргаффа). Получается, что последовательность одной цепочки предопределяет последовательность другой, и цепи как бы зеркально отражают друг друга. Такая закономерность, где нуклеотиды двух цепей располагаются упорядоченно, а также соединяются избирательно, называется принципом комплементарности. Кроме водородных соединений, двойная спираль взаимодействует и гидрофобно.
Две цепи разнонаправлены, то есть расположены в противоположных направлениях. Поэтому напротив трех"-конца одной находится пяти"-конец другой цепи.
Внешне напоминает винтовую лестницу, перилом которой является сахарофосфатный остов, а ступеньками — комплементарные основания азота.
Что такое рибонуклеиновая кислота?

РНК — это нуклеиновая кислота с мономерами, называющимися рибонуклеотидами.
По химическим свойствам она очень похожа на ДНК, так как обе являются полимерами нуклеотидов, представляющих собой фосфолированный N-гликозид, который выстроен на остатке пентозы (пятиуглеродного сахара), с фосфатной группой пятого углеродного атома и основания азота при первом углеродном атоме.
Она представляет собой одну полинуклеотидную цепочку (кроме вирусов), которая намного короче, чем у ДНК.
Один мономер РНК — это остатки следующих веществ:
- основания азота;
- пятиуглеродного моносахарида;
- кислоты фосфора.
РНК имеют пиримидиновые (урацил и цитозин) и пуриновые (аденин, гуанин) основания. Рибоза является моносахаридом нуклеотида РНК.
Отличия РНК и ДНК

Нуклеиновые кислоты отличаются друг от друга следующими свойствами:
- количество ее в клетке зависит от физиологического состояния, возраста и органной принадлежности;
- ДНК содержит углевод дезоксирибозу, а РНК — рибозу;
- азотистое основание у ДНК — тимин, а у РНК — урацил;
- классы выполняют различные функции, но синтезируются на матрице ДНК;
- ДНК состоит из двойной спирали, а РНК — из одинарной цепи;
- для нее нехарактерны действующие у ДНК;
- в РНК больше минорных оснований;
- цепи существенно отличаются по длине.
История изучения
Клетка РНК впервые была открыта биохимиком из Германии Р. Альтманом при исследовании дрожжевых клеток. В середине двадцатого века была доказана роль ДНК в генетике. Лишь тогда описали и типы РНК, функции и так далее. До 80-90% массы в клетке приходится на р-РНК, образующих совместно с белками рибосому и участвующих в биосинтезе белка.
В шестидесятых годах прошлого столетия впервые предположили, что должен существовать некий вид, который несет в себе генетическую информацию для синтеза белка. После этого научно установили, что есть такие информационные рибонуклеиновые кислоты, представляющие комплементарные копии генов. Их еще называют матричными РНК.
В декодировании записанной в них информации участвуют так называемые транспортные кислоты.
Позже стали разрабатываться способы выявления последовательности нуклеотидов и устанавливаться структура РНК в пространстве кислоты. Так было обнаружено, что некоторые из них, которые назвали рибозимами, могут расщеплять полирибонуклеотидные цепи. Вследствие этого стали предполагать, что в то время, когда зарождалась жизнь на планете, РНК действовала и без ДНК и белков. При этом все превращения производились с ее участием.
Строение молекулы рибонуклеиновой кислоты
Почти все РНК - это одиночные цепи полинуклеотидов, которые, в свою очередь, состоят из монорибонуклеотидов — пуриновых и пиримидиновых оснований.
Нуклеотиды обозначают начальными буквами оснований:
- аденина (А), А;
- гуанина (G), Г;
- цитозина (С), Ц;
- урацила (U), У.
Они связаны между собой трех- и пятифосфодиэфирными связями.

Самое разное количество нуклеотидов (от нескольких десятков до десятков тысяч) входит в строение РНК. Они могут формировать вторичную структуру, состоящую в основном из коротких двуцепочных тяжей, которые образовались комплементарными основаниями.
Структура молекулы рибнуклеиновой кислоты
Как уже было сказано, у молекулы имеется однонитевое строение. РНК получает вторичную структуру и форму в результате взаимодействия нуклеотидов между собой. Это полимер, мономером которого является нуклеотид, состоящий из сахара, остатка кислоты фосфора и основания азота. Внешне молекула похожа на одну из цепей ДНК. Нуклеотиды аденин и гуанин, входящие в состав РНК, относятся к пуриновым. Цитозин и урацил являются пиримидиновыми основаниями.
Процесс синтеза
Чтобы молекула РНК синтезировалась, матрицей является молекула ДНК. Бывает, правда, и обратный процесс, когда новые молекулы дезоксирибонуклеиновой кислоты образуются на матрице рибонуклеиновой. Такое встречается при репликации некоторых видов вирусов.
Основой для биосинтеза могут служить также другие молекулы рибонуклеиновой кислоты. В ее транскрипции, которая происходит в ядре клетки, участвуют много ферментов, но самым значимым из них является РНК-полимераза.
Виды
В зависимости от вида РНК, функции ее также отличаются. Существуют несколько видов:
- информационная и-РНК;
- рибосомальная р-РНК;
- транспортная т-РНК;
- минорная;
- рибозимы;
- вирусные.

Информационная рибонуклеиновая кислота
Такие молекулы еще называют матричными. Они составляют в клетке примерно два процента от всего количества. В клетках эукариот они синтезируются в ядрах на ДНК-матрицах, переходя затем в цитоплазму и связываясь с рибосомами. Далее, они становятся матрицами для синтеза белка: к ним присоединяются транспортные РНК, которые несут аминокислоты. Так происходит процесс преобразования информации, которая реализуется в уникальной структуре белка. В некоторых вирусных РНК она к тому же является хромосомой.
Жакоб и Мано являются открывателями этого вида. Не имея жесткой структуры, ее цепь образует изогнутые петли. Не работая, и-РНК собирается в складки и сворачивается в клубок, а в рабочем состоянии разворачивается.
и-РНК несет в себе информацию о последовательности аминокислот в белке, который синтезируется. Каждая аминокислота закодирована в определенном месте при помощи генетических кодов, которым свойственны:
- триплетность — из четырех мононуклеотидов возможно выстроить шестьдесят четыре кодона (генетического кода);
- неперекрещиваемость — информация движется в одном направлении;
- непрерывность — принцип работы сводится к тому, что одна и-РНК — один белок;
- универсальность — тот или иной вид аминокислоты кодируется у всех живых организмов одинаково;
- вырожденность — известными являются двадцать аминокислот, а кодонов — шестьдесят один, то есть они кодируются несколькими генетическими кодами.
Рибосомальная рибонуклеиновая кислота
Такие молекулы составляют подавляющее большинство клеточных РНК, а именно от восьмидесяти до девяноста процентов от общего количества. Они соединяются с белками и формируют рибосомы — это органоиды, выполняющие синтез белков.
Рибосомы состоят на шестьдесят пять процентов из р-РНК и на тридцать пять процентов из белка. Эта полинуклеотидная цепь без труда изгибается вместе с белком.
Рибосома состоит из аминокислотного и пептидного участков. Они расположены на контактирующих поверхностях.
Рибосомы свободно передвигаются нужных местах. Они не очень специфичны и могут не только считывать информацию с и-РНК, но и образовывать с ними матрицу.
Транспортная рибонуклеиновая кислота
т-РНК наиболее изучены. Они составляют десять процентов клеточной рибонуклеиновой кислоты. Эти виды РНК связываются с аминокислотами благодаря специальному ферменту и доставляются на рибосомы. При этом аминокислоты переносятся транспортными молекулами. Однако бывает, что аминокислоту кодируют разные кодоны. Тогда переносить их будут несколько транспортных РНК.
Она сворачивается в клубочек, когда неактивна, а функционируя, имеет вид клеверного листа.
В ней различаются следующие участки:
- акцепторный стебель, имеющий последовательность нуклеотидов АЦЦ;
- участок, служащий для присоединения к рибосоме;
- антикодон, кодирующий аминокислоту, которая присоединена к этой т-РНК.
Минорный вид рибонуклеиновой кислоты
Недавно виды РНК пополнились новым классом, так называемыми малыми РНК. Они, скорее всего, являются универсальными регуляторами, которые включают или выключают гены в эмбриональном развитии, а также контролируют процессы внутри клеток.
Рибозимы также недавно выявлены, они активно принимают участие, когда кислота РНК ферментируется, являясь при этом катализатором.
Вирусные виды кислот
Вирус способен содержать либо рибонуклеиновую кислоту, либо дезоксирибонуклеиновую. Поэтому с соответствующими молекулами они называются РНК-содержащими. При попадании в клетку такого вируса происходит обратная транскрипция — на базе рибонуклеиновой кислоты появляются новые ДНК, которые встраиваются в клетки, обеспечивая существование и размножение вируса. В другом случае происходит образование комплиментарной на поступившей РНК. Вирусы белков, жизнедеятельность и размножение идет без ДНК, а лишь на основе информации, содержащейся в РНК вируса.
Репликация
В целях улучшения общего понимания необходимо рассмотреть процесс репликации, в результате которого появляются две идентичные молекулы нуклеиновой кислоты. Так начинается деление клетки.
В ней участвуют ДНК-полимеразы, ДНК-зависимые, РНК-полимеразы и ДНК-лигазы.
Процесс репликации состоит из следующих этапов:
- деспирализация — происходит последовательное раскручивание материнской ДНК, захватывающей всю молекулу;
- разрыв водородных связей, при котором цепи расходятся, и появляется репликативная вилка;
- подстройка дНТФ к освободившимся основаниям материнских цепей;
- отщепление пирофосфатов от дНТФ молекул и образование фосфорнодиэфирных связей за счет выделяющейся энергии;
- респирализация.
После образования дочерней молекулы делится ядро, цитоплазма и остальное. Таким образом, образуются две дочерние клетки, полностью получившие всю генетическую информацию.
Кроме этого, кодируется первичная структура белков, которые в клетке синтезируются. ДНК в этом процессе принимает косвенное участие, а не прямое, заключающееся в том, что именно на ДНК происходит синтез, участвующих в образовании белков, РНК. Этот процесс получил название транскрипции.
Транскрипция
Синтез всех молекул происходит во время транскрипции, то есть переписывании генетической информации с определенного оперона ДНК. Процесс в некоторых моментах похож на репликацию, а в других существенно отличается от нее.
Сходствами являются следующие части:
- начало идет с деспирализации ДНК;
- происходит разрыв водородных связей между основаниями цепей;
- к ним комплементарно подстраиваются НТФ;
- происходит образование водородных связей.
Отличия от репликации:
- при транскрипции расплетается лишь участок ДНК, соответствующий транскриптону, в то время как при репликации расплетению подвергается вся молекула;
- при транскрипции подстраивающиеся НТФ содержат рибозу, и вместо тимина урацил;
- информация списывается лишь с определенного участка;
- после образования молекулы водородные связи и синтезированная цепь разрываются, а цепь соскальзывает с ДНК.
Для нормального функционирования первичная структура РНК должна состоять только из списанных с экзонов ДНК-участков.
У только что образованных РНК начинается процесс созревания. Молчащие участки вырезаются, а информативные сшиваются, образуя полинуклеотидную цепь. Далее, каждый вид имеет присущие только ему превращения.
В и-РНК происходит присоединение к начальному концу. К конечному участку присоединяется полиаденилат.
В т-РНК модифицируются основания, образуя минорные виды.
У р-РНК также метилируются отдельные основания.
Защищают от разрушения и улучшают транспортировку в цитоплазму белки. РНК в зрелом состоянии с ними соединяются.
Значение дезоксирибонуклеиновых и рибонуклеиновых кислот

Нуклеиновые кислоты имеют огромное значение в жизнедеятельности организмов. В них хранится, переносится в цитоплазму и передается по наследству дочерним клеткам информация о белках, синтезирующихся в каждой клетке. Они присутствуют во всех живых организмах, стабильность этих кислот играет важнейшую роль для нормального функционирования как клеток, так и всего организма. Любые изменения в их строении приведут к клеточным изменениям.
РНК - нуклеиновые кислоты, полимеры нуклеотидов, в состав которых входят остаток ортофосфорной кислоты, рибоза (в отличие от ДНК, содержащей дезоксирибозу) и азотистые основания - аденин, цитозин, гуанин и урацил (в отличие от ДНК, содержащей вместо урацила тимин). Эти молекулы содержатся в клетках всех живых организмов, а также в некоторых вирусах. РНК принимают участие во всех стадиях процесса генной экспрессии и биосинтеза белка
Структура и функции РНК.В цитоплазме клеток содержатся три основных вида РНК: матричная РНК (мРНК) выполняет роль матрицы при синтезе белка. Имеет сложную вторичную структуру. В целом линейная молекула мРНК имеет несколько двухспиральных шпилек, на концах которых располагаются «знаки» инициации и терминации трансляции транспортная РНК (тРНК) выполняет функцию посредника в ходе трансляции мРНК, транспортирует аминокислоту на рибосому. Вторичная структура имеет форму клеверного листа; рибосомная РНК (рРНК) формируют основу, с которой связываются специфические белки при образовании рибосомы.
Также в ядре клетки обнаружена ядерная РНК (яРНК), являющаяся предшественницей мРНК и тРНК.
Синтез РНК (транскрипция).1. Инициация. σ-Субединица РНК-полимеразы связывается с промотором – специальным участком ДНК. К σ-субединице присоединяется кофермент, цепи ДНК раскручиваются и образуется открытый транскрипционный комплекс. Первым нуклеотидом с 5’-конца является гуанин, который метилируется (кэпируется), эта стартовая точка гена называется кэп-сайт. 2. Элонгация. Происходит синтез комплементарной цепи. При этом пройденные участки из 15-20 нуклеотидов вновь спирализуются. 3. Терминация. Синтез РНК замедляется по достижении стоп-сигнала. Затем синтезируется цепь по коду дополнительного сигнала и еще 15 следующих за ним нуклеотидов. В процессе отделения РНК от матрицы экзонуклеаза отщепляет терминальные 15 нуклеотидов, а полиА-полимераза достраивает 150-200 полиА-нуклеотидов. 4. Процессинг РНК.
В отличие от ДНК, РНК не образуют двойных спиралей, но содержат короткие участки со спаренными основаниями. Это приводит к образованию субструктур, которые при двумерном изображении напоминают «шпильки» и петли, образующие фигуру типа «кленового листа». В таких структурах двухцепочечные участки соединены петлями.
Матричная РНК переносит генетическую информацию из клеточного ядра в цитоплазму. Так как мРНК считывается на рибосоме кодон за кодоном она не должна складываться в стабильную третичную структуру. Спариванию оснований препятствуют белки, ассоциированные с мРНК. Для мРНК характерно короткое время жизни, так как они быстро распадаются после трансляции. В сплайсинге предшественников мРНКпринимают участие малые ядерные РНК
Транспортные РНК участвуют в процессе трансляции в качестве промежуточного связующего звена между нуклеиновыми кислотами и белками. Это небольшие молекулы РНК из 70-90 нуклеотидов, которые с помощью своих антикодонов "узнают" за счет спаривания оснований определенные кодоны на мРНК.
По функциональному значению РНК делят на следующие виды:
1. транспортные РНК (тРНК) осуществляют кодирование аминокислот и перенос их в рибосомальный аппарат клетки в процессе биосинтеза белка. ТРНК локализованы в ядерном соке, митохондриях.
2. рибосомальные РНК (рРНК) являются структурной основой рибосом и выполняют в них разнообразные функции.
3. информационные или матричные РНК (мРНК) выполняют роль матрицы в процессе сборки полипептидных цепей в рибосомальном аппарате клетки.
4. вирусные РНК являются составными частями вирусных и фаговых рибонуклеопротеинов и несут всю информацию, необходимую для размножения вируса в клетках хозяина.
29. Рибосомы. Генетический код и его свойства. Трансляция.
Рибосома состоят из двух частей и представляют собой нуклеопротеины, состоящие из рРНК и белка в соотношении 1:1. Биологическая роль рРНК – являются структурной основой рибосом, взаимодействует с мРНК и тРНК в процессе биосинтеза белка, принимает участие в процессе сборки полипептидной цепи. У эукариот обнаружено 4 типа рРНК с различным коэф. седиментации: 18S(в малой части рибосомы), а 28S, 5,8S и 5S (сведбергов) – в большой части рибосомы.. Они различаются молекулярной массой (35 000-1 600 000) и локализацией в рибосомах. Вторичная структура рРНК характеризуется спирализацией цепи самой на себя, третичная – ее компактной укладкой. Рибосомы служат для биосинтеза белка из аминокислот по заданной матрице на основе генетической информации, предоставляемой матричной РНК (трансляция). Нередко с одной молекулой мРНК ассоциировано несколько рибосом, такая структура называется полирибосомой. Синтез рибосом у эукариот происходит в специальной внутриядерной структуре - ядрышке.
Генети́ческий код - свойственный всем живым организмам способ кодирования аминокислотной последовательности белков при помощи последовательности нуклеотидов. Информация о строении белка закодирована в ДНК с помощью генетического кода, который является линейным, непрерывным, триплетным, выражденным. Он является универсальным. В ДНК используется четыре нуклеотида - аденин (А), гуанин (G), цитозин (С), тимин (T). Эти буквы составляют алфавит генетического кода. В РНК используются те же нуклеотиды, за исключением тимина, который заменён похожим нуклеотидом - урацилом, который обозначается буквой U. В молекулах ДНК и РНК нуклеотиды выстраиваются в цепочки и, таким образом, получаются последовательности генетических букв. Для построения белков в природе используется 20 различных аминокислот. Каждый белок представляет собой цепочку или несколько цепочек аминокислот в строго определённой последовательности. Эта последовательность определяет строение белка, а, следовательно, все его биологические свойства. Набор аминокислот также универсален почти для всех живых организмов.Реализация генетической информации в живых клетках (то есть синтез белка) осуществляется при помощи двух матричных процессов: транскрипции (то есть синтеза мРНК на матрице ДНК) и трансляции генетического кода в аминокислотную последовательность (синтез полипептидной цепи на мРНК). Для кодирования 20 аминокислот, а также сигнала «стоп», означающего конец белковой последовательности, достаточно трёх последовательных нуклеотидов. Набор из трёх нуклеотидов называется триплетом. Принятые сокращения, соответствующие аминокислотам и кодонам.
Свойства:1Триплетность - значащей единицей кода является сочетание трёх нуклеотидов (триплет, или кодон).2Непрерывность - между триплетами нет знаков препинания, то есть информация считывается непрерывно.3Неперекрываемость - один и тот же нуклеотид не может входить одновременно в состав двух или более триплетов (не соблюдается для некоторых перекрывающихся генов вирусов, митохондрий и бактерий, которые кодируют несколько белков, считывающихся со сдвигом рамки).4Однозначность (специфичность) - определённый кодон соответствует только одной аминокислоте (однако, кодон UGA у Euplotes crassus кодирует две аминокислоты - цистеин и селеноцистеин)5Вырожденность (избыточность) - одной и той же аминокислоте может соответствовать несколько кодонов.7Универсальность - генетический код работает одинаково в организмах разного уровня сложности - от вирусов до человека (на этом основаны методы генной инженерии; есть ряд исключений, показанный в таблице раздела «Вариации стандартного генетического кода» ниже).8Помехоустойчивость - мутации замен нуклеотидов, не приводящие к смене класса кодируемой аминокислоты, называют консервативными; мутации замен нуклеотидов, приводящие к смене класса кодируемой аминокислоты, называют радикальными.
30. Регуляция синтеза белка у прокариотМутации, их виды и последствия.
Оперон – это функциональная единица ДНК прокариот. Строение:
1. Промотор – участок, к которому присоединяется фермент РНК-полимераза
2. Оператор – участок, которому присоединяется репрессор, который не дает РНК-полимеразе двигаться по ДНК
3. Структурные гены – содержат информацию о структуре белка. У прокариот в одном опероне находятся гены нескольких белков, необходимых для осуществления какой-либо биохимической реакции.
4. Терминатор – участок, в котором РНК-полимераза отсоединяется от ДНК.
Лактозный оперон открыли Жакоб, Моно и Львов в 1961 г. Его работа:
1. Когда в среде нет лактозы, кишечная палочка не вырабатывает ферменты, необходимые для ее расщепления, потому что к оператору присоединен репрессор, который не дает происходить транскрипции.
2. Когда в среде появляется лактоза, то она соединяется с белком-репрессором, он денатурирует и отсоединяется от оператора. Теперь ничто не мешает РНК-полимеразе делать иРНК, на которой рибосомы тут же делают белки.
3. Белки-ферменты расщепляют лактозу, в том числе и ту, что была присоединена к репрессору, он возвращается на место, транскрипция прекращается.
Мута́ция - стойкое (то есть такое, которое может быть унаследовано потомками данной клетки или организма) изменениегенотипа, происходящее под влиянием внешней или внутренней среды. Процесс возникновения мутаций получил название мутагенеза.
Спонтанные мутации возникают самопроизвольно на протяжении всей жизни организма в нормальных для него условиях окружающей среды
Индуцированными мутациями называют наследуемые изменения генома, возникающие в результате тех или иных мутагенных воздействий в искусственных (экспериментальных) условиях или при неблагоприятных воздействиях окружающей среды.
Существует несколько классификаций мутаций по различным критериям. В современной учебной литературе используется и более формальная классификация, основанная на характере изменения структуры отдельных генов, хромосом и генома в целом. В рамках этой классификации различают следующие виды мутаций:
геномные; хромосомные; генные.
Геномные : - полиплоидизация (образование организмов или клеток, геном которых представлен более чем двумя (3n, 4n, 6n и т. д.) наборами хромосом) и анеуплоидия (гетероплоидия) - изменение числа хромосом, не кратное гаплоидному набору
При хромосомных мутациях происходят крупные перестройки структуры отдельных хромосом. В этом случае наблюдаются потеря (делеция) или удвоение части (дупликация) генетического материала одной или нескольких хромосом, изменение ориентации сегментов хромосом в отдельных хромосомах (инверсия), а также перенос части генетического материала с одной хромосомы на другую (транслокация)
На генном уровне изменения первичной структуры ДНК генов под действием мутаций менее значительны, чем при хромосомных мутациях, однако генные мутации встречаются более часто. В результате генных мутаций происходят замены, делеции и вставки одного или нескольких нуклеотидов, транслокации, дупликации и инверсии различных частей гена. В том случае, когда под действием мутации изменяется лишь один нуклеотид, говорят о точечных мутациях.
Возможны четыре генетических последствия точковых мутаций: 1) сохранение смысла кодона из-за вырожденности генетического кода (синонимическая замена нуклеотида), 2) изменение смысла кодона, приводящее к замене аминокислоты в соответствующем месте полипептидной цепи (миссенс-мутация), 3) образование бессмысленного кодона с преждевременной терминацией (нонсенс-мутация). В генетическом коде имеются три бессмысленных кодона: амбер - UAG, охр - UAA и опал - UGA
4) обратная замена (стоп-кодона на смысловой кодон).
31.Ферментативный гидролиз белков.Протеолитические ферменты, их специфичность, активация.
Переваривание белков пищи начинается в желудке. Здесь в кислой среде белок денатурирует и подвергается действию пепсина – эндонуклеазы желудочного сока, которая гидролизует внутренние связи, образованные карбоксильной группой ароматических аминокислот (Phe +, Tyr+, Thr+).
Образовавшиеся полинуклеотиды поступают в просвет тонкого кишечника. Здесь на них действуют четыре фермента панкреатического секрета: трипсин, химотрипсин, эластаза и карбоксипептидаза. Трипсин и химотрипсин гидролизуют связи, образованные аминогруппами основных (+Lys, +Arg) и ароматических аминокислот (+Phe, +Tyr, +Thr). Эластаза – эндонуклеаза широкого спектра действия. Карбоксипептидаза – экзонуклеаза, гидролизующая связь С-концевой аминокислоты.+N-CH(R)-COOH
В пристеночной области кишечника олигонуклеотиды гидролизуются до свободных аминокислот ферментами кишечного сока – аминопептидазой, ди- и трипептидазой. Образовавшиеся свободные аминокислоты всасываются и поступают в кровоток.
ПРОТЕОЛИТИЧЕСКИЕ ФЕРМЕНТЫ (протеазы), ферменты класса гидролаз, катализирующие гидролиз (протео-лиз) пептидных связей. Место расщепления пептидной связи в полипептидной цепи определяется позиционной и субстратной специфичностью фермента и пространств. структурой гидролизуемого субстрата (белка или пептида).
Различают экзопептидазы, расщепляющие связи вблизи С- или N-конца цепи (соотв. карбоксипептидазы и аминопеп-тидазы)и эндопептидазы (протеиназы), гидролизующие связи, удаленные от концевых остатков (напр., трипсин). Лишь ограниченное число протеолитических ферментов обладает строгой субстратной специфичностью. К ним относят, напр., ренин, гидроли-зующий связь между остатками лейцина в положениях 10 и 11 в ангиотензиногене (предшественник ангиотензина пептида, участвующего в регуляции кровяного давления).
В зависимости от локализации фермента протеолиз происходит при разл. рН. Так, протеолитические ферменты желудка (напр., пепсин, гастриксин) функционируют при рН 1,5-2, лизосомные ферменты-при рН 4-5, а протеолитические ферменты сыворотки крови, тонкого кишечника и др.-при нейтральных или слабощелочных значениях рН. Нек-рые протеолитические ферменты используют в качестве кофактора ионы металлов-Са2+, Mg2+ и др.
Дефектные и чужеродные белки деградируют в клетке при участии АТФ-зависимой системы протеолиза. У эукариот (все организмы, кроме бактерий и синезеленых водорослей) эта система включает низкомол. белок убикитин, образующий с белками-субстратами конъюгат, и протеазы, расщепляющие этот конъюгат.
32.Общая схема источников и расходования аминокислот в организме. Незаменимые аминокислоты. Общие пути катаболизма аминокислот.
Доставленные с кровотоком аминокислоты в печень имеют несколько основных путей метаболизма:
1. Биосинтеза белка – протекает как в самой печени, так и в других тканях.
2. Трансаминирование и использование углеродного скелета для биосинтеза глюкозы, жирных кислот и кетоновых тел, биосинтеза АТФ.
3. Декарбоксилирование с образованием биологически активных биогенных аминов (дофамин, норадреналин и адреналин (синтезируются изначально из аминокислоты тирозина), серотонин, мелатонин и триптамин (синтезируются из триптофана).
4. Биосинтез нуклеотидов, порфиринов, гормонов.
Трансаминирование и дезаминирование аминокислот
В процессе трансаминирования аминогруппа переносится на кетокислоту, в роли которой выступает 2-кетоглутарат, пируват или оксалоацетат, которые образуют глутамат, аланин и аспартат соответственно. Далее аминогруппа аланина и аспартата переносится на 2-кетоглутарат с образованием глутамата (непрямое дезаминирование). Глутамат – единственная аминокислота подвергающаяся окислительному дезаминированию.
Глюкогенные аминокислоты образуют одно из пяти соединений, которые через оксалоацетат вовлекаются в глюконеогенез. Кетогенные аминокислоты образуют ацетоацетат или ацетил-КоА. Ряд аминокислот дают метаболиты обоих видов и являются одновременно кетогенныи и глюкогенными.
Декарбоксилирование аминокислот
В результате отщепления карбоксильной группы аминокислоты образуют биогенные амины, многие из которых выполняют важные биологические функции.
Так, серин образует этанолмаин, включаемый в состав фосфолипидов; глутамат – нейромедиатор γ-аминомасляную кислоту (ГАМК); гистидин – медиатор гистамин.
Дезактивация биогенных аминов происходит путем их окислительного дезаменирования и дальнейшего окисления в карбоновые кислоты.
Двенадцать из двадцати аминокислот могут быть синтезированы de novo в организме. При этом цистеин и тирозин образуются из незаменимых метионина и фенилаланина, а аргинин и гистидин синтезируются в незначительных количествах. Оставшиеся восемь получаются в результате переноса аминогруппы глутамата на кетокислоты-предшественники
реакция в обмене аминок-т 11рис
Вещества, образованные при внутриклеточном гидролизе или проникающие в клетку из крови, на втором этапе обычно превращаются в пировиноградную кислоту, ацетильную группу (в составе ацетил-S-КоА) и в некоторые другие мелкие органические молекулы. Локализация второго этапа – цитозоль и митохондрии. Часть энергии рассеивается в виде тепла и примерно 13% энергии вещества усваивается, т.е. запасается в виде макроэргических связей АТФ Ацетил-SКоА включается в реакции цикла трикарбоновых кислот и окисляется до углекислого газа. Выделенные атомы водорода соединяются с НАД и ФАД и восстанавливают их. После этого НАДН и ФАДН2 переносят водород в цепь дыхательных ферментов, расположенную на внутренней мембране митохондрий. Здесь в результате процесса под названием "окислительное фосфорилирование" образуется вода и главный продукт биологического окисления – АТФ.
33.ПЕРЕАМИНИРОВАНИЕ (трансаминирование), обратимый перенос аминогруппы из молекулы одного орг. соед. в молекулу другого . Наиб. роль переаминирование играет в биохимии в процессах метаболизма азотистых оснований. Переаминирование аминокислот может происходить и вне клетки в присут. пиридоксаль-5"-фосфата, однако скорость р-ции в 106 раз меньше.
Часть аминокислот, поступивших в избыточном количестве с пищей, либо образовавшиеся в результате распада тканевых белков, теряют аминогруппу и превращаются в кетокислоту.
В процессе трансаминирования (рис.10) аминогруппа переносится на кетокислоту, в роли которой выступает 2-кетоглутарат, пируват или оксалоацетат, которые образуют глутамат, аланин и аспартат соответственно. Далее аминогруппа аланина и аспартата переносится на 2-кетоглутарат с образованием глутамата (непрямое дезаминирование). Глутамат – единственная аминокислота подвергающаяся окислительному дезаминированию.
В результате дезаминирования аминогруппа отщепляется в виде аммиака, который метаболизируется печенью.
Углеродные скелеты аминокислот могут быть вовлечены в ЦТК и окислены до конечных продуктов, либо использованы для биосинтеза глюкозы (глюкогенные аминокислоты), жирных кислот или кетоновых тел (кетогенные аминокислоты).
Глюкогенные аминокислоты образуют одно из пяти соединений, которые через оксалоацетат вовлекаются в глюконеогенез.
Кетогенные аминокислоты образуют ацетоацетат или ацетил-КоА. Ряд аминокислот дают метаболиты обоих видов и являются одновременно кетогенныи и глюкогенными (пролин, лейцин и др).
Нарушение нормального течения переаминирования в организме наблюдается при патологич. состояниях, напр. при инфаркте миокарда, заболеваниях печени. Контроль концентрации аминотрансфераз используется в этих случаях как ср-во диагностики.
ДЕЗАМИНИРОВАНИЕ (деаминирование), удаление группы NH2 из молекул орг. соединений. Сопровождается замещением аминогруппы на др. группу или образованием кратной связи. Осуществляется под действием HNO2, нитрозилсерной к-ты, N2O3, N2O4, H2O, H2, а также ферментов.
34. Образование аммиака. Транспорт аммиака.Восстановительное аминирование. Амиды и их физиологическое значение.
БИОГЕННЫЕ АМИНЫ Образуются при декарбоксилировании аминокислот. Биогенные амины (БА) являются непептидными азотсодержащими производными аминокислот. К ним относятся гормоны надпочечников (адреналин, норадреналин), щитовидной железы (тироксин, трийодтиронин), медиаторы ЦНС (ацетилхолин, ГАМК, дофамин), медиатор воспаления (гистамин), этаоламин, аминопропанол и др. Некоторые из этих соединений являются составными частями других биомолекул.
![]()
Амины, образовавшиеся при декарбоксилировании аминокислот, часто являются биоактивными веществами.
Серотонин – нейромедиатор проводящих путей. Образуется в надпочечниках и ЦНС.
Аминомасляная кислота (ГАМК). В нервных клетках декарбоксилирование глутамата приводит к образованию ГАМК, которая служит основным тормозным медиатором высших отделов мозга.
Аминомасляная кислота (ГАМК). В нервных клетках декарбоксилирование глутамата приводит к образованию ГАМК, которая служит основным тормозным медиатором высших отделов мозга.
Гистамин выполняет в организме человека следующие функции: --стимулирует секрецию желудочного сока, слюны; --повышает проницаемость капилляров, вызывает отеки, снижает АД; --сокращает гладкую мускулатуру легких, вызывает удушье; --участвует в формировании воспалительной реакции – вызывает расширение сосудов, покраснение кожи, отечность ткани; – вызывает аллергическую реакцию; -выполняет роль нейромедиатора; -является медиатором боли.

реакция в обмене аминок-т рис13
Восстановительное аминирование - это превращение кетонов в соответствующие им амины.
35. Особенности обмена отдельных аминокислот и их роль в образовании важнейших биологически активных веществ.
СПЕЦИФИЧЕСКИЕ ПУТИ ОБМЕНА ОТДЕЛЬНЫХ АМИНОКИСЛОТ. Эти пути обмена определяются различиями в строении радикалов аминокислот, поэтому они разнообразны и многочисленны. Вступая в эти специфические реакции, АК-ты принимают участие во многих важных процессах:а) в синтезе гормонов и нейромедиаторов, б) в синтезе простетических групп сложных белков - хромопротеинов и нуклеопротеинов, в) в синтезе сложных липидов, г) синтезе веществ, содержащих макроэргическую связь и являющихся источником энергии для клеток, д) в обезвреживании токсических веществ.
ОБМЕН ГЛИЦИНА И СЕРИНА.Это заменимые аинокислоты, которые превращаются друг в друга.Основным путем распада глицина является его распад на СО2, Н2О и метилен-ТГФК (активный С1):
Активный С1, образовавшийся из глицина участвует в синтезе пуриновых азотистых оснований. Кроме того, глицин всей своей молекулой участвует в синтезе гема гемоглобина и других гемопротеинов, в синтезе пуриновых азотистых оснований, в синтезе парных желчных кислот (гликохолевая кислота), в синтезе креатина, в синтезе трипептида глютатиона. Также глицин в печени участвует в обезвреживании бензойной кислоты, которая превращается в гиппуровую кислоту:
ОБМЕН СЕРУСОДЕРЖАЩИХ АМИНОКИСЛОТ: МЕТИОНИНА И ЦИСТЕИНА.Метионин - это незаменимая аминокислота, а цистеин - заменимая.S-аденозил-метионин участвует в реакциях трансметилирования. Наиболее важный из них синтез фосфатидилхолина из фосфатидилэтаноламина, обезвреживание биогенных аминов с участием О-метилтрансфераз, синтез адреналина из норадреналина, синтез ацетилхолина из холина и Ацетил-КоА, синтез креатина, который в виде креатинфосфата является резервной формой макроэргических связей и участвует в обеспечении нервной ткани и работающей мышцы АТФ.
Синтез креатина. В синтезе креатина участвуют и другие аминокислоты - аргинин и глицин. В почках из аргинина и глицина образуется гуанидинацетат, который метилируется в печени с участием S-аденозил-метионина и в результате образуется креатин:
Гомоцистеин участвует в синтезе амикислот - цистеина (гомоцистеин + серин). Креатин подвергается фосфорилированию с участием АТФ, в результате образуется соединение с макроэргической связью - креатинфосфат. Это обратимая реакция, которая катализируется ферментом креатинфосфокиназой (КФК).
ОБМЕН ЦИСТЕИНА.Это заменимая аминокислота, она синтезируется из серина, гидроксильная группа которого замещается SН-группой, которую поставляет гомоцистеин. Цистеин в составе белков-ферментов своей -SН группой участвует в образовании каталитического центра (тиоловые протеазы), а также участвует в образовании дисульфидных связей, которые принимают участие в формировании третичной и четвертичной структуры белков. Также цистеин необходим для синтеза трипептида глютатиона, который состоит из цистеина и глютаминовой кислоты. Молекула глутатиона условно обозначается как (Г-SН). Глютатион способен легко окисляться и восстанавливаться:Окисляясь, глютатион предохраняет от окисления другие вещества, например, двухвалентное железо гемоглобина в эритроцитах:Цистеин подвергается и распаду, при этом он окисляется и декарбоксилируется, в результате образуется таурин, который участвует в образовании парных желчных кислот (таурохолевая и др.) в печени. Серная кислота, которая образуется из таурина, участвует в обезвреживании токсических веществ в печени.
ОБМЕН АРГИНИНА Аргинин - частично незаменимая аминокислота. Она образуется в ходе синтеза мочевины в печени из карбомоилфосфата при участии аспарагиновой кислоты и орнитина. Аргинин участвует в синтезе креатина в почках, являясь донором гуанидиновой группы в образовании гуанидинацетата. В составе белков аргинин как полярная положительно заряженная аминокислота участвует в образовании ионных связей и в формировании гидратной оболочки белков.
КОНКРЕТНАЯ РОЛЬ КАЖДОЙ АМИНОКИСЛОТЫ:Глутаминовая кислотаа) подвергается прямому окислительному дезаминированиюс образованием альфа-кетоглутарата,б) вступает в реакции трансаминирования, которые катализируют специфические трансаминазы,в) является субстратом для синтеза глютамина, который является транспортной формой аммиака и участвует в синтезе мочевины в печени, также глютамин участвует в синтезе пуриновых оснований нуклеотидов и нуклеиновых кислот, аминосахаров и аминопроизводных липидов.г) принимает участие в косвенном дезаминировании АК-т,д) участвует в синтезе трипептида глютатиона,е) является субстратом для образования гамма- аминомасляной кислоты.
Аспарагиновая кислотаа) участвует в реакциях трансаминирования,б) в синтезе мочевины, как донор NН2-группы,в) в синтезе пиримидиновых оснований (всей молекулой) и как донор NН2-группы - в синтезе пуриновых оснований,г) в синтезе аспарагина.
Обе аминокислоты участвуют в формировании третичной и четвертичной структур белков, так как являются полярными заряженными аминокислотами и в образовании гидратной оболочки белков, а также эти аминокислоты принимают участие в формировании активных центров ферментов. Амидирование аспарагиновой и глутаминовой кислот приводит к образованию глутамина и аспарагина, необходимых для синтеза белков.
ОБМЕН ЦИКЛИЧЕСКИХ АМИНОКИСЛОТ ФЕНИЛАЛАНИНА И ТИРОЗИНАФенилаланин является незаменимой аминокислотой, а тирозин - заменимая аминокислота.Тирозин вступает в многочисленные реакции в различных тканях. В результате этих превращений тирозин не только распадается до конечных продуктов, но и дает промежуточные метаболиты, из которых образуются ряд важных соединений, некоторые из которых являются биологически активными веществами. Из тирозина образуются:а) гормоны мозгового слоя надпочечников адреналин и норадреналин,б) меланины - пигменты кожи, волос, радужной оболочки глаза,в) йодсодержащие гормоны щитовидной железы - тироксин и трийодтиронин.
фенилкетонурия - нарушен синтез фенилаланин-гидроксилазы, поэтому фенилаланин превращается в фенилпируват, который оказывает токсическое воздействие на развитие некоторых отделов головного мозга.2) альбинизм - нарушен синтез ферментов, превращающих ДОФА в ДОФА-хром, поэтому нарушается синтез меланинов. 3) алкаптонурия - нарушен синтез диоксигеназы гомогентизиновой кислоты, она выделяется с мочой, моча приобретает черный цвет.4) кретинизм - нарушен синтез йодиназы, что приводит к нарушению синтеза йодсодержащих гормонов щитовидной железы.5) может быть нарушен синтез фермента тирозиназы, который катализирует превращение тирозина в ДОФА, следовательно будет нарушаться синтез гормонов мозгового слоя надпочечников и меланина.Из всех этих заболеваний в настоящее время удается лечить фенилкетонурию, для этого из рациона ребенка исключают фенилаланин и увеличивают в пище количество тирозина. Если ребенка держать на этой диете до 6-7 лет, тогда не возникает умственная отсталость, т.к. к 6-7 годам успевают развиться отделы головного мозга, развитие которых задерживается при избытке в ткани мозга фенилпирувата.
36.Биосинтез мочевины. Орнитиновый цикл мочевинообразования.
Образующийся в результате реакций дезаминирования аммиак является токсичным для организма.  Его дезактивация с образованием нетоксичных мочевины и креатина происходит в печени, куда он предварительно должен быть транспортирован кровью. Трансортными формами аммиака являются аспарагин и глутамин, образующиеся амидированием аспартата и глутамата. Реакции катализируются аспарагин- и глутаминсинтетазой. В печени аспарагиназа и глутаминаза отщепляют амидную группу в виде аммиака.
Его дезактивация с образованием нетоксичных мочевины и креатина происходит в печени, куда он предварительно должен быть транспортирован кровью. Трансортными формами аммиака являются аспарагин и глутамин, образующиеся амидированием аспартата и глутамата. Реакции катализируются аспарагин- и глутаминсинтетазой. В печени аспарагиназа и глутаминаза отщепляют амидную группу в виде аммиака.
В печени аммиак карбоксилируется с образованием карбамоилфосфата, который вовлекается в орнитиновый цикл. На последнем этапе этого процесса образуется аргинин, который распадается на орнитин и мочевину, либо преобразуется в креатин. В мышцах креатин фосфорилируется и используется в качестве донора фосфатной группы при регенерировании АТФ из АДФ. Образовавшиеся креатинин и мочевина выводятся из организма.
В карте обмена АК-т от аммиака до карбамоил-фосфата к мочеине.